Hacia un ecosistema micorrícico del aprendizaje en la era de los modelos de lenguaje
Los modelos de lenguaje no son un nodo más en la red del aprendizaje. Son la red filamentosa subterránea que reconfigura todo el ecosistema. Por qué la metáfora del grafo conectivista envejeció y qué viene después.

Por qué la metáfora del grafo conectivista envejeció y qué viene después
Preludio biográfico · 14 años después
Me crié en los páramos de León. Paisaje austero, encinares dispersos, suelos donde el secreto está siempre debajo. De niño me apasionaba descifrar los signos de la naturaleza: las formas que se repiten sin parecerse, las redes invisibles que sostienen a los árboles, los hongos que aparecen donde menos se esperan. Aquella mirada nunca se fue.
Décadas después, el 31 de diciembre de 2012, último día de aquel año, publiqué en este blog un artículo sobre redes de micorrizas, simbiosis y comunidades de aprendizaje. Trabajaba con metáforas bioinspiradas, leía a Wiley, observaba el comportamiento parasitario que empezaba a dominar las redes sociales digitales y especulaba con que las comunidades de aprendizaje podían pensarse como ecosistemas simbióticos antes que como grafos de información. No supe entonces que aquella intuición, anclada en una infancia de hongos y raíces, me esperaría catorce años después con la fuerza de una hipótesis confirmada.
Los modelos de lenguaje a gran escala han introducido en el ecosistema educativo una capa que en 2012 era impensable, y la metáfora micorrícica resulta hoy más precisa que el día en que la propuse. Lo que sigue es la actualización de aquel texto, escrita con la ventaja de los años y con la incomodidad de descubrir que el problema del parasitismo, lejos de resolverse, ha encontrado terreno fresco donde extenderse.
Hay textos que uno escribe sin saber que está dejando una pista para el yo futuro. El páramo leonés siempre supo algo que solo ahora empiezo a articular del todo.
La imagen que ya no nos sirve
Vuelvo del páramo al despacho universitario y son las dos imágenes que cargo desde hace meses. La primera es la que dibujábamos en pizarras hace quince años para explicar el conectivismo: círculos que representan personas, recursos e instituciones, líneas que los unen, flechas que indican circulación de información. Un grafo limpio, plano, geométrico. La segunda es la que sigo viendo cuando salgo a pasear las arboledas al lado del rio en León en primavera (por contextualizar en la época del año que estamos): árboles visibles que parecen organismos individuales pero que, según sabemos hoy, están conectados bajo tierra por una red filamentosa que los mantiene vivos. La primera imagen es nítida y didáctica. La segunda es turbia y verdadera.
Sostengo que la primera imagen ya no nos sirve para nombrar lo que ocurre cuando un estudiante universitario aprende en 2026. No porque sea falsa, sino porque es insuficiente. Hay algo en la segunda imagen que la primera no puede albergar. Y ese algo coincide, sospechosamente, con lo que la irrupción de los modelos de lenguaje a gran escala ha hecho con nuestras prácticas docentes. Lo que sigue es una propuesta de cambio de metáfora, con todo lo que un cambio de metáfora arrastra.
El grafo conectivista y su techo conceptual
El conectivismo nació, hace exactamente dos décadas, para nombrar algo que las teorías de aprendizaje anteriores no podían acomodar (Siemens, 2005). El cognitivismo seguía pensando el aprendizaje como almacenamiento individual de representaciones mentales. El constructivismo lo entendía como construcción interna de esquemas. Ambos enfoques colocaban al sujeto en el centro y trataban la red de información como un entorno externo del que el sujeto extrae materiales. El conectivismo desplazó ese centro. Propuso que el aprendizaje, en la era digital, ya no se explica adecuadamente sin atender a la red de conexiones que el sujeto consigue construir, mantener y atravesar. El conocimiento, en esta formulación, ya no reside íntegramente en la cabeza del aprendiz, sino en la red.
Fue una propuesta valiente y oportuna. Permitió pensar lo que las teorías anteriores no veían: que aprender con Wikipedia, con redes profesionales en línea, con repositorios digitales y con colegas geográficamente dispersos no era simplemente "lo de siempre con tecnología". Era otra cosa. Y necesitaba marco.
Sin embargo, conviene señalar que la metáfora subyacente al conectivismo es computacional. El grafo, el nodo, la conexión, el flujo de información. Es vocabulario heredado de la teoría de redes y de la informática. Esto no es un defecto en sí mismo. Es una decisión metafórica que tenía sentido en un momento histórico en el que internet emergía como infraestructura dominante. Pero toda metáfora ilumina ciertas cosas y oscurece otras. Y la metáfora del grafo, conviene reconocerlo, oscurece varias dimensiones del aprendizaje que hoy resultan críticas.
Primero, los grafos no tienen subsuelo. Sus nodos están todos en un mismo plano, son todos del mismo tipo lógico, son todos visibles e identificables. El grafo no sabe representar lo que está debajo de los nodos sosteniéndolos. Y todo lo que los modelos de lenguaje hacen ocurre, precisamente, en una capa que el grafo conectivista no podía dibujar.
Segundo, los grafos no tienen ciclos estacionales. No respiran. No pasan por épocas de crecimiento, descanso, latencia o renovación. Su topología es estática. El aprendizaje universitario, en cambio, está atravesado por ritmos académicos, vitales, profesionales. La metáfora del bosque sabe esto. La del grafo, no.
Tercero, los grafos no tienen patógenos. No conciben la enfermedad. En un grafo, una conexión funciona o no funciona, está activa o inactiva. En un ecosistema, en cambio, las conexiones pueden estar colonizadas por organismos oportunistas, pueden estar transmitiendo señales tóxicas, pueden estar al servicio del parásito en lugar del huésped. Volveremos sobre esto cuando hablemos de autoridad sintética.
Cuarto, y quizá más importante, los grafos no tienen simbiosis obligada. Cada nodo del conectivismo es, en principio, autosuficiente. Puede existir aunque otros nodos desaparezcan. Un ecosistema, en cambio, es un tejido donde algunos organismos solo existen porque otros también existen. Las micorrizas son uno de los ejemplos más estudiados: ni el hongo ni la planta sobreviven plenamente sin el otro. Y la formación universitaria, sostengo, se parece mucho más a la simbiosis obligada que a la cooperación opcional entre nodos autónomos.
El conectivismo necesita, no actualización, sino traducción a otra metáfora. Y esa metáfora ya existe, hace décadas que se trabaja en biología, y converge con tradiciones pedagógicas latinoamericanas y peninsulares que llevan tiempo pensando lo educativo como ecosistema. Vamos a ella.
La metáfora micorrícica como giro epistémico
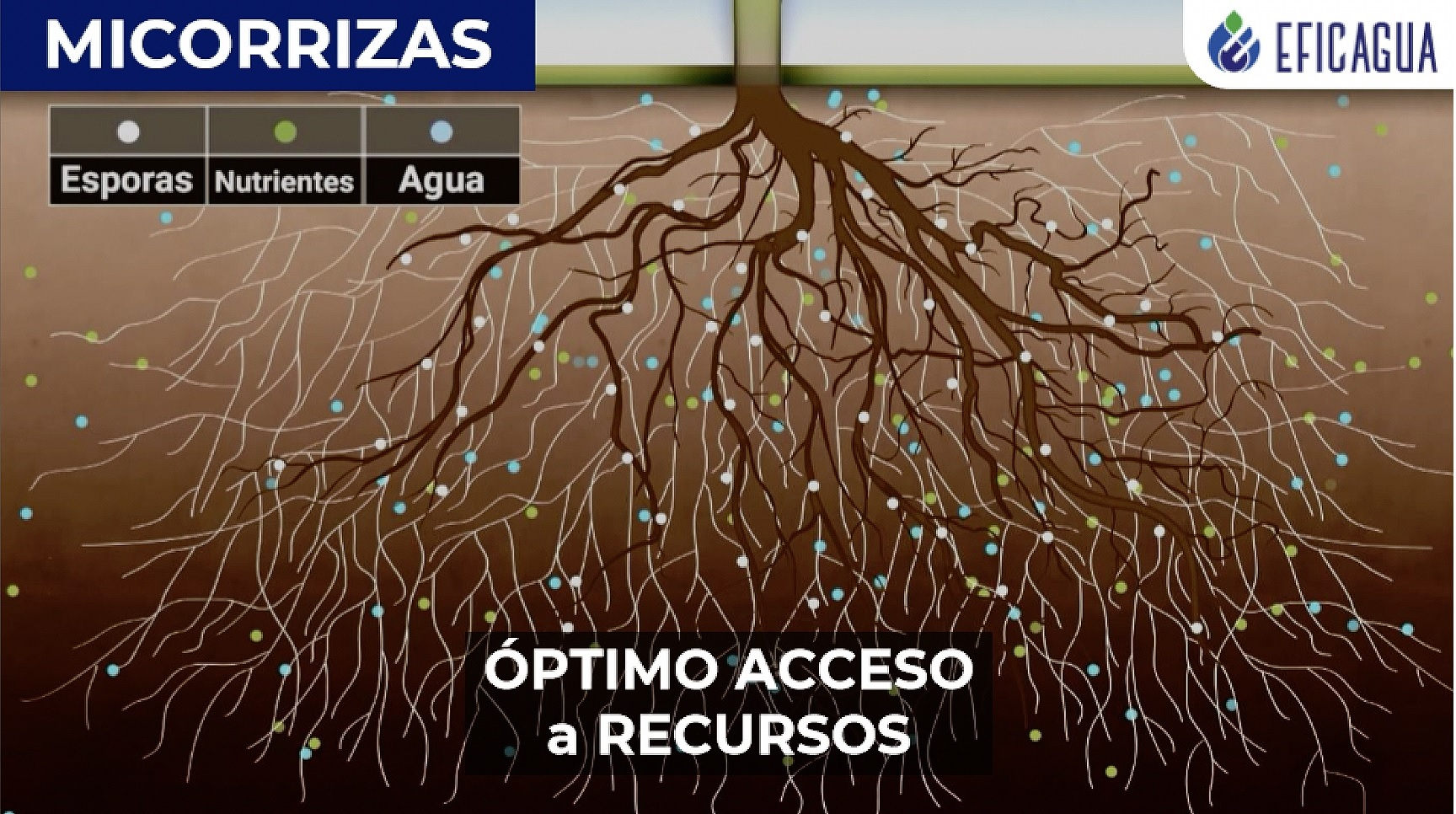
Las micorrizas son asociaciones simbióticas entre las raíces de las plantas y determinados hongos del suelo. La planta proporciona al hongo azúcares fabricados mediante fotosíntesis. El hongo proporciona a la planta agua y nutrientes que sus filamentos pueden alcanzar en volúmenes de suelo que las raíces, por sí solas, jamás explorarían. La asociación es tan extendida que aproximadamente el ochenta por ciento de las plantas terrestres dependen de ella en algún grado. Cuando hablamos de bosques, lo que vemos son árboles. Lo que sostiene a los árboles, por debajo, es una red filamentosa de hifas fúngicas que conecta organismos aparentemente separados y por la que circulan agua, nutrientes, señales químicas y, según algunas hipótesis, también información compleja.
Suzanne Simard, ecóloga forestal de la University of British Columbia, popularizó con su libro Finding the Mother Tree (2021) la idea de que los bosques son comunidades cooperativas atravesadas por estas redes, a las que la revista Nature había bautizado en 1997 como wood wide web. Conviene introducir aquí una anotación académica: parte de las afirmaciones más fuertes de Simard, especialmente las que sugieren formas de comunicación intencional entre árboles, han sido criticadas por colegas botánicos en Trends in Plant Science, con réplica posterior. No estamos ante un consenso cerrado. Pero el núcleo biológico, la existencia y la importancia de las redes micorrícicas, es ciencia firme. Y es ese núcleo el que nos interesa metafóricamente.
¿Qué transfiere la metáfora micorrícica a nuestra discusión sobre el aprendizaje?
Transfiere, primero, la idea de infraestructura invisible estructurante. Lo visible (el árbol, el estudiante, el docente) no se sostiene por sí mismo. Está atravesado por una capa que no vemos pero que decide buena parte de lo que ocurre arriba. Si la red micorrícica funciona, el bosque funciona. Si la red se rompe, los árboles pueden seguir en pie, pero pierden capacidad de adaptación, de recuperación, de comunicación. Aplicado al aprendizaje contemporáneo, los modelos de lenguaje son justamente esa capa invisible. No porque sean ocultos, sino porque su funcionamiento estructural está por debajo de lo que el grafo conectivista podía representar.
Transfiere, segundo, la idea de simbiosis obligada. La micorriza no es una opción para la planta. Es, en la mayoría de los casos, una condición de existencia. Esto no significa que cualquier asociación sea buena. Significa que la asociación, sea buena o mala, es constitutiva. Trasladado al aula universitaria, los modelos de lenguaje ya no son una herramienta opcional que el docente decide introducir. Son una capa con la que el estudiante interactúa quiera o no el docente. Pretender prohibirlos institucionalmente es como pretender prohibir el subsuelo. Lo que cabe es decidir qué tipo de simbiosis cultivamos con ellos.
Transfiere, tercero, la idea de distribución no proporcional de recursos. En una red micorrícica, los nutrientes no fluyen igualmente entre todos los participantes. Algunos árboles dan más de lo que reciben. Otros reciben más de lo que dan. La red contiene desigualdades estructurales que la metáfora del grafo simétrico tendía a borrar. Trasladado a nuestro caso, el flujo entre estudiante, modelo de lenguaje y plataforma es asimétrico por construcción. Reconocerlo es el primer paso para no quedar atrapado en él.
Transfiere, cuarto, la idea de comunicación sin proximidad. Árboles que crecen lejos físicamente pueden estar conectados por hifas comunes. La proximidad no es condición de la conexión. Esta propiedad, banal en biología, tiene implicaciones interesantes para pensar la formación universitaria distribuida y las comunidades de aprendizaje que se sostienen sin contacto presencial.
Tenemos, entonces, una metáfora con cuatro propiedades transferibles que el grafo conectivista no podía representar. Veamos ahora qué aspecto tiene el ecosistema completo cuando lo dibujamos.
Componentes del ecosistema
El siguiente esquema representa una sección transversal del ecosistema micorrícico del aprendizaje. Conviene leerlo de arriba hacia abajo, prestando atención tanto a lo visible como a lo subterráneo.

🖼️ IMAGEN 1: Sección transversal del ecosistema. Tres árboles (nodo humano en teal, nodo de contenido en navy, nodo institucional en amber) en la superficie. Línea de suelo cognitivo. Red filamentosa coral subterránea conectando las raíces. Caja central etiquetada "Red micorrícica · modelos de lenguaje". Pie: "Sustrato generativo: corpus, datos, infraestructura técnica".
Recorramos sus componentes en orden, prestando atención a las funciones pedagógicas que cumple cada uno.
Los árboles visibles son los tres tipos de nodo que el conectivismo clásico ya reconocía. Lo que cambia respecto a la formulación original no es su existencia, sino su estatus relativo. Los árboles ya no son la totalidad del ecosistema. Son su parte emergida, su superficie. Una facultad universitaria observada desde el conectivismo clásico parecería un conjunto de árboles aislados: docentes, contenidos, plataformas. Observada desde el ecosistema micorrícico, esa misma facultad aparece como un bosque cuyo subsuelo cuenta tanto o más que su dosel.
El suelo cognitivo es la interfaz entre lo visible y lo subterráneo. Está compuesto por los datos curriculares, los diseños instruccionales, las plataformas que median la interacción y las prácticas docentes que decantan en hábitos. Es la zona donde lo institucional se encuentra con lo orgánico, donde los nutrientes circulan y donde los problemas de compactación empiezan a notarse antes de que los árboles muestren síntomas.
La red micorrícica es donde se sitúan los modelos de lenguaje. No los sitúo como un cuarto árbol al lado de los tres anteriores, sino como infraestructura filamentosa subterránea que conecta las raíces de los tres tipos clásicos y que circula recursos entre ellos. Esta es la diferencia conceptual más importante respecto a planteamientos anteriores. Los modelos de lenguaje no son ni docentes sintéticos, ni recursos mejorados, ni plataformas convencionales. Son la red que reconfigura la circulación de información entre los tres tipos. Y, como cualquier red micorrícica, pueden funcionar mutualísticamente o pueden estar colonizados por organismos oportunistas.
El clima académico son las políticas educativas, las culturas institucionales y las orientaciones estratégicas que regulan las condiciones de posibilidad del ecosistema. Determinan qué se permite, qué se incentiva, qué se penaliza. Un mismo bosque puede prosperar o degradarse según el clima que lo rodea. Una misma facultad puede convertir los modelos de lenguaje en aliados pedagógicos o en parásitos extractivos según el clima académico que la institución sostenga.
Las raíces son los aprendices. Son la interfaz orgánica entre los árboles y la red. Es a través de las raíces como un estudiante accede a la red micorrícica, le pide nutrientes y le entrega azúcares (atención, datos, fragmentos de su propia producción). Esta es una zona delicada: la salud del estudiante depende de la salud de su sistema radicular y de la red que lo nutre, en proporciones que la pedagogía debería tomarse en serio.
Los frutos y semillas son los contenidos producidos en la interacción: ensayos, trabajos finales, presentaciones, debates. En el conectivismo clásico, estos contenidos se entendían como producción individual del estudiante. En el ecosistema micorrícico, son producto compuesto del árbol y de la red. Reconocerlo abre el debate sobre autoría y evaluación que la universidad está apenas empezando a sostener con seriedad.
Tipologías de simbiosis y su traducción pedagógica
La biología distingue, desde hace más de un siglo, tres formas básicas de relación simbiótica. Cada una tiene una traducción pedagógica precisa cuando se aplica al ecosistema que estamos describiendo.

El mutualismo describe una relación en la que ambas partes obtienen recursos que no podrían producir solas. Aplicado al aprendizaje con modelos de lenguaje, el mutualismo aparece cuando el estudiante aporta criterio, contexto y dirección, y el modelo aporta articulación, alcance, formulaciones alternativas y velocidad de exploración. El resultado es un contenido que ninguno de los dos habría producido por separado, y un aprendiz que ha refinado su capacidad de juicio precisamente porque ha tenido que ejercerla en interacción con un agente que le devuelve resultados que necesitan validación. Es el escenario al que apuntan, con razón, autores como Mollick (2024) cuando hablan de cointeligencia. Pero el mutualismo no es la condición por defecto. Es una conquista pedagógica que requiere diseño.
El comensalismo describe una relación en la que una parte se beneficia y la otra no pierde ni gana. En nuestro contexto, esto ocurre cuando el estudiante usa el modelo para tareas auxiliares (resumir un texto largo, generar variaciones de un párrafo, traducir un fragmento) sin que el uso afecte sustantivamente su trayectoria formativa. Es una zona pedagógicamente neutra: ni educativamente valiosa ni problemática. La mayoría del uso cotidiano de modelos de lenguaje en aulas universitarias probablemente cae aquí. No es lo que nos preocupa.
El parasitismo es el modo problemático. Una parte extrae recursos del otro sin devolver beneficio equivalente. Tres modalidades merecen distinguirse. La primera es el parasitismo del estudiante sobre la red, que ocurre cuando se delega en el modelo la totalidad de la tarea cognitiva (redactar el trabajo, generar el análisis, decidir la postura argumental) y el estudiante recibe a cambio una pieza terminada que finge propia. Hay extracción de recursos, no hay devolución pedagógica, no hay aprendizaje. La segunda modalidad es el parasitismo de la plataforma sobre el estudiante, que ocurre cuando los datos generados por la interacción son extraídos y reciclados sin que el estudiante reciba a cambio una mejora proporcional de su experiencia formativa. La tercera, quizá la más invisible, es el parasitismo de la red sobre los nodos clásicos, que ocurre cuando la mediación de los modelos atrofia la consulta a docentes, libros y comunidades, generando dependencia funcional y empobrecimiento del ecosistema en conjunto.
En 2012, al estudiar otro ecosistema digital, observábamos que la proporción de parasitismo frente a mutualismo en redes sociales era aproximadamente de nueve a uno (Santamaría, 2012b). No tenemos motivos sólidos para suponer que la proporción será mejor en el ecosistema micorrícico del aprendizaje sin intervención pedagógica deliberada. Lo que cabe esperar, sin diseño, es la misma asimetría.
Patógenos del ecosistema cognitivo
La metáfora micorrícica tiene una ventaja epistémica adicional respecto al grafo conectivista. Permite pensar patologías del ecosistema, no solo nodos y conexiones. Un grafo no se enferma. Un bosque sí. Y esta diferencia conceptual abre un vocabulario crítico que nos hace falta.

En esta imagen 3 identifico cuatro patógenos que merecen nombre propio y vigilancia pedagógica.
El primero es la autoridad sintética sin verificación. Los modelos de lenguaje producen afirmaciones con coherencia gramatical y articulación retórica que generan, en quien las recibe, un efecto subjetivo análogo al de la autoridad fundada. Pero su autoridad no procede de experiencia situada, de revisión por pares ni de tradición institucional. Procede de minimización estadística de error sobre un corpus. Bender y colaboradoras (2021) describieron este problema con la metáfora del loro estocástico, que aunque polémica capta una intuición correcta: la fluidez gramatical no implica validación epistémica. El patógeno no consiste en que los modelos produzcan errores. Consiste en que los producen con la misma elegancia retórica con que producen aciertos, y la única defensa es la verificación activa, que muchos estudiantes no realizan porque no han sido enseñados a realizarla.
El segundo patógeno es el monocultivo cognitivo. Cuando una sola red domina la circulación de recursos, los demás nodos del ecosistema se atrofian. En agricultura sabemos hace décadas que el monocultivo aumenta los rendimientos a corto plazo y reduce la resiliencia del sistema a largo plazo. En lo cognitivo ocurre algo análogo. Si el estudiante deja de consultar libros, deja de hablar con docentes, deja de frecuentar comunidades, y canaliza toda su consulta a través de la red micorrícica, la red se vuelve más útil de manera inmediata y el ecosistema se vuelve más frágil de manera estructural. Williamson (2017) había advertido, antes de la era de los grandes modelos, sobre los riesgos de la datificación educativa cuando una sola lógica domina la organización del conocimiento. El monocultivo cognitivo es la versión actualizada de aquella advertencia.
El tercer patógeno es la compactación del suelo. En agricultura, el paso continuo de maquinaria pesada compacta el suelo, reduce su porosidad, dificulta la circulación de agua y la respiración de las raíces. En lo educativo, la compactación ocurre cuando las plataformas técnicas, las exigencias administrativas y los protocolos extractivos saturan el espacio entre los componentes del ecosistema. El estudiante deja de tener tiempo para divagar, para aburrirse, para tropezar productivamente con un libro inesperado. El docente deja de tener margen para diseñar fuera del estándar. La red, atrapada en una matriz compactada, pierde capacidad de generar variaciones interesantes.
El cuarto patógeno son las especies oportunistas. En un ecosistema sano, ciertas especies oportunistas cumplen funciones útiles. En un ecosistema estresado, esas mismas especies pueden invadir nichos y desplazar a otras. En el ecosistema cognitivo, las especies oportunistas son las que prosperan precisamente donde los nodos clásicos se han debilitado: contenido generado masivamente sin curaduría, plataformas de microaprendizaje sin diseño pedagógico, agentes conversacionales orientados a la fidelización antes que al aprendizaje. No son malignas por naturaleza. Son problemáticas cuando proliferan sin contrapeso.
Pedagogía del ecosistema
¿Qué hacemos con todo esto? La pregunta es legítima, y necesita respuesta operativa, no solo conceptual.
Sostengo, primero, que la respuesta NO es sustituir. Sustituir docentes por agentes, libros por respuestas generadas, comunidades por hilos de conversación con modelos, equivale a clarear el bosque para que crezca solo la red micorrícica. Es un error ecológico. La red existe gracias a los árboles, y desaparece sin ellos.
Sostengo, segundo, que la respuesta tampoco es prohibir. Pretender que la red micorrícica no exista equivale a hacer agricultura ignorando el subsuelo. Funciona durante un tiempo, hasta que el suelo se agota.
La respuesta que propongo es la cultivación. Cultivar un ecosistema es diseñar las condiciones bajo las cuales los componentes prosperan en relaciones mutualistas, los patógenos se mantienen bajo control y el clima académico permite ciclos sostenibles. Cultivación no es sinónimo de control ni de naturalización pasiva. Es trabajo deliberado, situado y paciente.

La pluralidad de nodos es el contrapeso al monocultivo. Diseñar pedagógicamente para que cada microunidad de aprendizaje exija contacto con al menos dos tipos distintos de nodo, no solo con la red. Leer un libro entero, conversar con un colega, frecuentar un archivo. No por nostalgia institucional, sino por salud ecosistémica. Hug (2007) sostenía ya que el microaprendizaje tenía sentido solo si se articulaba con otras dimensiones más extensas. Lo mismo vale aquí, ampliado.
La alfabetización en trazabilidad es el contrapeso a la autoridad sintética sin verificación. Convertir la verificación de fuentes, la triangulación con otros nodos y el reconocimiento de los límites del modelo en competencia transversal de la universidad. No como contenido optativo de talleres puntuales, sino como práctica integrada en toda asignatura que use modelos de lenguaje. Si no enseñamos esto, expondremos a las próximas promociones a una opacidad disfrazada de claridad expositiva.
La comunidad como humedad es el contrapeso al simulacro social. La metáfora biológica es exacta: un suelo seco no permite la circulación de nutrientes en la red micorrícica. Una formación universitaria sin comunidad real, sin docencia con cuerpo, sin pares con quienes pensar en voz alta, sin tutorías presenciales o virtuales auténticas, deja a la red micorrícica trabajando sobre suelo seco. Y la red, por sofisticada que sea, no puede sustituir lo que no se le encomendó hacer. Wenger (1998) y antes Lave y Wenger (1991) ya habían argumentado que el aprendizaje vive en comunidades de práctica. Esta intuición no envejece, gana vigencia.
Los ciclos de descanso son el contrapeso a la compactación del suelo. Toda ecología productiva sostenible exige ritmos. Un bosque sin descanso se agota. Un currículo sin pausas reflexivas, sin tiempos para sedimentar, sin ventanas para que el estudiante haga sin producir, sin ratos de ocio académico productivo, se compacta. La pedagogía del ecosistema necesita reivindicar el descanso, no como lujo extracurricular, sino como condición de salud del sistema.
Estos cuatro principios no son recetas cerradas. Son orientaciones de diseño. Cada universidad, cada facultad, cada asignatura tendrá que traducirlos a sus condiciones específicas. Lo importante no es el listado, sino la lógica subyacente: pasamos de pensar el aprendizaje como conexión de nodos a pensarlo como cultivo de un ecosistema, con todo lo que la palabra cultivo implica de paciencia, de saberes situados y de respeto por los tiempos largos.
El clima no se decreta, se cultiva
Llevamos un trecho del recorrido y conviene mirar atrás antes de cerrar. Hemos sostenido que el conectivismo clásico, valioso en su momento, opera con una metáfora computacional del grafo que ya no alcanza para nombrar el aprendizaje contemporáneo. Hemos propuesto que la metáfora micorrícica, anclada en biología, ofrece cuatro propiedades transferibles que el grafo no podía representar: infraestructura invisible estructurante, simbiosis obligada, distribución no proporcional de recursos, comunicación sin proximidad. Hemos descrito los componentes de un ecosistema en el que los modelos de lenguaje no son un cuarto árbol, sino la red filamentosa subterránea que reconfigura la circulación entre los tres árboles visibles. Hemos distinguido tres tipos de simbiosis y cuatro patógenos. Y hemos propuesto cuatro principios operativos de cultivación.
Quedan, por supuesto, preguntas sin respuesta. La metáfora micorrícica, ¿no antropomorfiza ahora a los modelos del modo opuesto, asignándoles un papel orgánico que no les corresponde? Conviene tomarse la objeción en serio. Lo que defendemos no es que los modelos sean organismos vivos, sino que cumplen, en el ecosistema del aprendizaje, una función estructural análoga a la de las redes micorrícicas en los bosques. Es una analogía operativa, no una identificación ontológica. Puede revisarse, puede afinarse, puede sustituirse. Lo que no parece sostenible es seguir pensando el ecosistema actual con la metáfora del grafo plano.
Otra pregunta legítima: si los modelos son la red micorrícica, ¿quién decide sobre la red? Aquí la metáfora deja de funcionar y conviene reconocerlo. Las redes micorrícicas reales no tienen propietarios. Las que hoy median el aprendizaje universitario sí. Empresas privadas, en su mayoría angloparlantes, con intereses comerciales legítimos pero no necesariamente alineados con los pedagógicos. Este desajuste entre la metáfora ecológica y la realidad económica del ecosistema actual es una de las tensiones más serias de las que tendremos que ocuparnos en los próximos años, y posiblemente sea el lugar donde la conversación pedagógica deba encontrarse con la conversación política y regulatoria.
El clima académico no se decreta desde un rectorado, no se diseña en un decreto ministerial, no se delega en un departamento de innovación docente. Se cultiva en miles de pequeñas decisiones que cada docente, cada coordinador de asignatura, cada estudiante toman cada semana. Es trabajo invisible y acumulativo, mucho más parecido al cuidado de un bosque que al diseño de una infraestructura. Y exige, sobre todo, paciencia con los tiempos que no son los del semestre lectivo.
Termino con una pregunta que me llevo puesta y que dejo abierta para quien siga leyendo. Si el ecosistema micorrícico del aprendizaje es lo que ya tenemos, y si el clima en el que se desarrolla lo cultivamos colectivamente decisión a decisión, ¿qué tipo de bosque estamos sembrando en las universidades hispanohablantes para los próximos veinte años? La respuesta, sospecho, dependerá menos de la tecnología disponible y más de la calidad pedagógica de las preguntas que sepamos hacernos antes de que el clima se decante por su cuenta.
Referencias
Bender, E. M., Gebru, T., McMillan-Major, A., y Shmitchell, S. (2021). On the dangers of stochastic parrots: Can language models be too big? En Proceedings of the 2021 ACM Conference on Fairness, Accountability, and Transparency (pp. 610-623). Association for Computing Machinery. https://doi.org/10.1145/3442188.3445922
Hug, T. (Ed.). (2007). Didactics of microlearning: Concepts, discourses and examples. Waxmann.
Lave, J., y Wenger, E. (1991). Situated learning: Legitimate peripheral participation. Cambridge University Press.
Mollick, E. (2024). Co-intelligence: Living and working with AI. Portfolio.
Santamaría, F. (2012a, 12 de agosto). Microlearning - microaprendizaje: Definiciones y características [Entrada de blog]. Blog de Fernando Santamaría. https://fernandosantamaria.com/microlearning-microaprendizaje-definiciones-y-caracteristicas/
Santamaría, F. (2012b, 31 de diciembre). Redes de micorrizas, simbiosis y tipos de asociaciones para las comunidades [Entrada de blog]. Blog de Fernando Santamaría. https://fernandosantamaria.com/redes-de-micorrizas-simbiosis-y-tipos-de-asociaciones-para-las-comunidades/
Siemens, G. (2005). Connectivism: A learning theory for the digital age. International Journal of Instructional Technology and Distance Learning, 2(1), 3-10.
Siemens, G. (2007). Connectivism: Creating a learning ecology in distributed environments. En T. Hug (Ed.), Didactics of microlearning: Concepts, discourses and examples. Waxmann.
Simard, S. (2021). Finding the Mother Tree: Discovering the wisdom of the forest. Knopf.
Wenger, E. (1998). Communities of practice: Learning, meaning, and identity. Cambridge University Press.
Williamson, B. (2017). Big data in education: The digital future of learning, policy and practice. Sage.